The Cannabis Breeder's Bible: ГЛАВА 17 - ЭВОЛЮЦИОННЫЙ КАНАЛ КОНОПЛИ
The Cannabis Breeder's Bible - Greg GreenГЛАВА 17: ЭВОЛЮЦИОННЫЙ КАНАЛ КОНОПЛИ
Гарантии способности растений конопли к самовоспроизведению
Когда мы имеем дело с неслучайным скрещиванием между родственными особями растений, в том числе и самоопылением (так же называемым автогамией, в отличие от нормального скрещивания отдельного мужского и отдельного женского растения, называемого аллогамией), мы можем увидеть в популяции проблемы, связанные с качеством потомства. Дарвин обратил внимание, что потомство при родственном скрещивании было не столь жизнеспособным, как при неродственном скрещивании. В природе практически не попадаются популяции гермафродитов конопли, в естественных условиях они могут быть найдены лишь в некоторых регионах. Из-за того, что двудомные растения более распространены, мы предполагаем, что гермафродитизм не подходит для большинства ареалов, где обнаружена конопля. Но это предположение необходимо обсудить, потому что близкородственное скрещивание может сделать сорт более устойчивым к специфическим условиям среды, и человек участвует в распространении и гермафродитов, и двудомных растений. Небольшие популяции гермафродитной конопли по всему миру похоже доказывают, что этот признак не совсем неподходящий для семейства конопли.
В 1941 году Рональд Алимер Фишер предположил «преимущество автоматической передачи». С его трудами можно ознакомиться в Интернете по адресу http://www.library.adelaide.edu.au/ digitised/fisher/index.html.
Фишер открыл, что в процессе опыления происходит автоматический преимущественный отбор. Самоопылённое растение, являясь гермафродитом, даёт будущему потомству и пыльцу и яйцеклетки. В популяции, где самоопыления не происходит, мужские растения отдают пыльцу женским растениям. Самоопылённые растения, однако, будут содержать аллель, обуславливающую гермафродитный признак, и Фишер обнаружил, что эта аллель при передаче имеет преимущество в соотношении 3:2 по сравнению с не самоопыляемыми растениями. Это означает, что самоопыленные растения обычно несут эту аллель дальше в популяцию, пока не достигается равновесие, и ген не исчезает из генофонда. Короче, самоопыляемые растения, похоже, подвергаются естественному отбору, чтобы оставаться гермафродитами и передавать этот признак потомству, однако это не значит, что самоопыляемая популяция гермафродитов не может вдруг стать двудомной популяцией, растения в которой не самоопыляются.
Семена конопли довольно тяжёлые. Хотя они могут переноситься на далёкие дистанции животными и птицами, в большинстве случаев они прорастают в непосредственной близости от материнского растения, с которого упали. В этом виновата гравитация. Подавляющее большинство материнских растений конопли заканчивает свою жизнь до созревания потомства, так что вероятность того, что произойдёт естественное обратное, скрещивание очень мала. Однако если мамка выживет, шансы на беккросс в дикой природе сильно повышаются.
Мы знаем, что конопля противодействует этому, потому что это однолетнее растение и большинство сортов конопли демонстрируют признаки, предотвращающие обратное скрещивание. Иногда растение не опыляется собственной пыльцой или семена в нём не вызревают. Это называется «гомоморфной несовместимостью» или «защитная самонесовместимость».
Антезис (от греч. ánthesis - цветение) это период, когда цветок раскрывается или становится сексуально-функциональным – время образования шишки и развития цветов. Мужские и женские растения демонстрируют чёткую разницу во времени цветения. Мужские растения развиваются быстрее женских, обычно даже до того, как девочки начинают цвести. Гермафродит же становится таким чаще всего уже в фазу цветения женского растения. Если до появления гермафродитов в популяции уже появилась пыльца с других мужских растений, она, как более доступная, скорее всего раньше будет использована для опыления. Опылённые растения очень редко превращаются в гермафродитов после получения пыльцы, если только не подвергаются очень сильным стрессам
Так каковы преимущества, ради которых эволюция сохранила признак гермафродитизма? Они сводятся к вариациям в сортах, повышающих шансы на адаптацию и приспособляемость потомства.
Если вам нужна гарантия на то, что воспроизводство произойдёт, вы получите её посредством самоопыления. Вы, возможно, задаётесь вопросом: если сорт без проблем соответствует окружающим условиям, почему бы ему не заняться размножением посредством самоопыления? Самоопыление скорее подразумевается как процесс самосохранения в стрессовых условиях, когда растение, пребывающее в кризисе, вынуждено опылить себя. Но даже в нормальных условиях, разве не в интересах растения было бы воспроизводиться автогамно? Мы вновь возвращаемся к той же проблеме. Может, гермафродитизм это крайняя точка и при хороших, и при плохих условиях? Это говорит нам, что данная система скрещивания тесно связана с родственным скрещиванием.
Так как при родственном скрещивании происходит угнетение (о котором мы сейчас скажем) различных возможностей и способности растения к адаптации, мы можем сравнить это с потомством того же вида, которое не участвует в автогамии или процессах выборочного размножения. Эта формула выражается как:
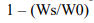
Ws это измеренная выживаемость самоопылённого потомства, а W0 это измеренная выживаемость не самоопылённого потомства. Выживаемость измеряется признаками на каждой стадии развития растения и их сочетаемостью с условиями среды. Причины угнетения могут быть обнаружены у сортов с большим количеством гомозиготных признаков в популяции.
Лучше всего угнетение при близкородственном скрещивании прослеживается у генетически однородных сортов. Если два несвязанных чистых сорта встречаются, они производят первое поколение гибридов F1, которые имеют некоторые характеристики выживаемости в популяции. Гетерозис это тенденция, проявляющаяся у гибридов, появление в потомстве качеств, превосходящих аналогичные качества обоих родителей. Это так же называется «гибридная сила».
Инбридинг, близкородственное скрещивание, повышает гомозиготность, а неродственное скрещивание, кроссбридинг, увеличивает гетерозиготность. При кроссбридинге гибриды получаются сильнее, результатом чего является ускорение роста и плодовитости. Разница между обоими родителями в результате даёт силу гибридам. В большой популяции, где скрещиваются не близкородственные сорта, в потомстве первого поколения проявляются смешанные черты обоих родителей и иногда проявляются доминантные фенотипы обоих сортов. Возможности популяции заключаются в различии в частоте генов, встречающихся в ней. Мы рассмотрим эти смешанные характеристики и то, как они сочетаются и производят сильных гибридов прямо сейчас.
Жизненная сила гибридов
Гетерозис это критерий различий в частоте генов, умноженный преобладающими отклонениями. Эта гипотеза утверждает, что гетерозис вызван преобладающим проявлением «вредных генов» при усиливающейся гомозиготности и гетерозиготность исчезает при слиянии соответствующих гомозиготных признаков. Можно так же предположить, что гетерозис происходит из-за уничтожения в гетерозиготном поколении F1 вредных рецессивных признаков, вносимых одним из родителей при скрещивании. Всё это станет более понятным через секунду, когда мы рассмотрим возможные комбинации генов в популяции применимо к нашей модели увеличения силы гибридов.
Итак, что такое вредоносное рецессивное состояние? Вредоносные аллели уменьшают вероятность выживания индивида, тогда как безопасные (не вредоносные) аллели повышают шансы на выживание индивида (это плохие и хорошие признаки, влияющие на выживаемость растения). Если рецессивный признак является ослабляющим, то растения с этим признаком произведут меньшее потомство, которое привнесёт этот признак в популяцию. Растения с вредоносными аллелями обычно погибают раньше здоровых особей.
Вредоносные аллели существуют и преумножаются в неоднородной популяции из-за фактора неслучайного скрещивания. Вредоносный рецессивный признак вообще заставляет растение использовать больше ресурсов для компенсации проблемы. Иногда вредоносный рецессивный признак заставляет растение расти выше с более длинными междоузлиями, что позволяет ему получить больше света и подавить меньшие растения, у которых нет вредоносной аллели. Итак, в итоге вредоносный ген подавляет не вредоносный ген посредством других доминирующих признаков растения. Однако это довольно редкое явление и в большинстве случаев число вредоносных аллелей остаётся небольшим из-за процесса естественного отбора.
Селекционеры могут сохранить растения, которые бы не выжили в дикой природе из-за их вредоносных генов. Это зависит от того, что мы называем вредоносным геном. Плохая сопротивляемость определённым типам паразитов может считаться вредоносным признаком, но IBL сорта, выращиваемые в закрытом помещении, могут и не подвергнуться атаке этих паразитов и этот признак останется незамеченным. Если бы это растение существовало в местности, где присутствовали бы эти паразиты, вредоносный признак ослабевал бы популяцию и, скорее всего не сохранился бы в ней. Вредоносная аллель обычно обременяет растение в популяции, и оно использует больше энергии и ресурсов для выживания. Оно использует больше ресурсов, чем растения без вредоносных генов несущих этот признак, таким образом, считается, что оно убыточнее в сравнении с ними. Но растения с вредоносными признаками могут доказать свою успешность в борьбе с проблемами, с которыми они столкнулись. И опять же, наличие вариаций повышает адаптивность. Поэтому рецессивные аллели не классифицируются как вредоносные или дающие преимущества. То же относится и к доминантным аллелям.
Если мы поймём это и вспомним, что мы узнали о рекомбинации генетического материала конопли, мы сможем понять силу гибридов в модели, приведённой ниже.
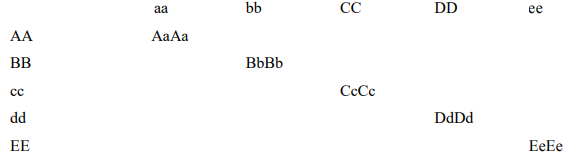
Основной элемент тут это растение с генотипом AaBbCcDdEe. Обратите внимание, что ни у кого из родителей не встречается такая комбинация. Генотипы родителей AABBccddEE и aabbCCDDee. У первого родителя, AABBccddEE, два рецессивных гена (сс и dd). У второго родителя, aabbCCDDee, три рецессивных гена (aa, bb, ee). У потомства генотип AaBbCcDdEe, то есть доминантный по всем пяти признакам A, B, C, D и E. Все эти аллели А В С D E присутствуют в потомстве, и их доминантность выразится в его соответствующих фенотипах, что даёт растению его гибридную силу. Это потомство уникально! Пожалуйста помните, тем не менее, что мы используем эту модель в контексте повышения силы гибридов с рецессивными или частично рецессивными аллелями*. Теперь вы увидели, как в популяции посредством естественного отбора гибридов может появиться превосходящее родителей по своим параметрам растение. Вот почему важно растить большие популяции для отбора того самого особого материнского растения, которое лучше остальных. Обычно это особое материнское растение является сильным гибридом.
* Комбинация доминантных и рецессивных признаков может дать тот особый сильный гибрид.
Есть масса противоречий на счёт силы гибридов, потому что в этом есть и обратное явление, когда доминантная аллель не является лучшей для соответствия условиям среды. При помощи случайного скрещивания вы в конкретной окружающей среде должны помочь усилению черт, подходящих для этой среды, с последующим удалением вредоносных аллелей, не подходящих для неё. Сила и приспособляемость при скрещивании гибридов F1 и в дальнейших поколениях будет уменьшаться.
«Сверхдоминантность» это термин, так же используемый для описания силы гибридов. Настоящая сверхдоминантность происходит, когда существуют многочисленные аллели одного и того же признака, то есть их больше двух. Если происходит выборочное скрещивание, по одному и тому же признаку можно произвести исключительное множество гибридов. Как вы можете догадаться, чем больше создаётся гибридов, тем больше произойдёт рекомбинаций и тем выше шансы на получение сильного растения, подходящего для конкретной среды обитания. Короче говоря, селекционеры развивают и улучшают генетику конопли, перемешивая генетические коды, а не совершая близкородственные скрещивания. Близкородственное скрещивание стабилизирует сорт, закрепляя гомозиготные признаки.
Предисловие к половой эволюции конопли
Самоопылённые растения обычно производят больше семян из-за давления отбора. Наличие доступной и готовой пыльцы это часть гарантии воспроизводства.
В 1932 году Джон Бердон Халдейн опубликовал книгу «Причины эволюции». Халдейн сделал открытие, что существует высокая вероятность закрепления полезных для адаптации аллелей при самоопылении, нежели при внешних скрещиваниях, потому что они закрепляются быстрее, чем при других видах селекции. Этот процесс, известный как решето Халдейна, так же несёт негативный эффект, потому что долгосрочные преимущества теряются в пользу краткосрочных. Это термин известен как «адаптированность» и впервые использован С.К. Джейном в предложенной им гипотезе адаптированности в публикации «Эволюция при близкородственном скрещивания у растений» в 1976 году.
В 1957 году доктор наук Г. Ледьярд Стеббинс опубликовал труды «Регулярность преобразований в цветке», «Самооплодотворение и популяционная изменчивость высших растений» и «Нежизнеспособность, слабость и стерильность межвидовых гибридов» среди прочих важных трудов того времени. Стеббинс предполагает, что эволюция самоопыления это объединение предварительных гарантий воспроизведения и адаптивности. Стеббинс открыл, что самоопыление даёт преимущество видам для сохранения хорошо адаптированных генотипов в течении длительный периодов времени, если окружающая среда подходит популяции.
Мы рассмотрели преимущества и недостатки самоопыления. Вкратце, самоопыление увеличивает гарантии на воспроизведение вида, но может уменьшить долговременные преимущества (Стоит отметить, что здесь мы не принимаем во внимание, что целью гровера является получение не опылённой травы. Здесь мы рассматриваем эволюционные процессы двудомной и однодомной конопли). В своей публикации «Эволюция самооплодотворения и угнетение при родственном скрещивании» 1985 года, Р. Ланде и Д. В. Шемске открыли, что фактически в дикой популяции растений присутствует беспорядочный отбор по степени самоопыляемости, что препятствует широкому распространению гермафродитизма. И самоопыление (автогамия) и обычное размножение (аллогамия) это виды скрещивания и они зависят от совместимости растений. Самооплодотворение в популяции, где в основном происходит внешнее скрещивание, становится причиной высокой степени угнетения, из-за большей частоты рецессивных гомозиготных генотипов, которого не происходит при неродственном скрещивании. Таким образом, близкородственное скрещивание в популяциях с преобладающим внешним скрещиванием помогает развиваться механизмам неродственного скрещивания, а не самоопыления.
В 1996 году М. Шируп и Ф.Б. Кристинсен в своей публикации «Межродственное и неродственное скрещивание у растений» и в 1997 году М. Фишер и Д. Мэттис в своей публикации «Структура скрещивания и угнетение при близкородственном и неродственном скрещивании у редкого растения Gentianella Germanica (Gentianaceae)» предположили, что если мы рассмотрим в основном самоопыляющиеся популяции, то увидим, что у растений хорошо выражены их сильные стороны, способствующие адаптации. Скрещивание с другими растениями, даже из другой популяции, могут стать причиной «угнетения при неродственном скрещивании», и потомство в этом скрещивании будет слабее, чем потомство при самоопылении. Таким образом, степень полезности самоопыления зависит от того, подходит ли растение к конкретным условиям среды.
Близкородственное скрещивание может повлиять на механизмы опыления и это называется «эволюция механизмов опыления» и может быть прослежено в различиях между однодомными и двудомными типами растений. Двудомные растения, похоже, развились из однодомных растений.
Развитие двудомных растений это процесс, протекающий в два шага. Первый шаг в развитии двудомного растения это «мутация, приводящая к стерильности мужского растения», которое перестаёт производить пыльцу. Это состояние известно как гинодиэция и единственные функционирующие репродуктивные органы это женские соцветия. В таком типе популяций присутствуют женские растения со стерильными мужскими цветами и гермафродиты. Ген мужской стерильности распространится сразу же, как только попадёт в популяцию.
Андродиэция это аналогичное состояние, но в нём у растения присутсвуют мужские цветы и стерильные женские цветы, а так же цветы-гермафродиты, но это редкое явление.
Следующий шаг в развитии двудомности это мутация, затрагивающая производство пыльцы, происходящая на растениях без гинодиэции. Растения попытаются компенсировать гинодиецию, увеличивая количество мужских особей и уменьшая производство женских. Поскольку происходит модификация, уменьшается производство яйцеклеток у гермафродитов, а не женских растений. Если выживают гермафродиты с усиленной мужской функцией, этот ген распространится в популяции. В результате в потомстве будут женские растения (с нефункционирующими мужскими органами) и мужские растения (с нефункционирующими женскими органами). По прошествии времени эти растения разовьют или мужские или женские половые органы и разделятся на два отдельных пола, то есть станут двудомными. Это очень интересно, не правда ли? Это так же объясняет, почему наследование пола и выражение пола у конопли происходит так сложно.
Чистые линии несут адаптивные ценности, которые могут быть замечены и в природе и в лаборатории. Близкородственное скрещивание может дать хорошие результаты в популяции, в которой существует высокая степень соответствия, потому что они больше подходят для среды, в которой выведены.
Самоопыление имеет репродуктивные преимущества и так же схоже с адаптивными ценностями чистых линий. Хотя близкородственное скрещивание ведёт к угнетению, это угнетение проявляется, только если сорт попадает в неблагоприятную среду или равновесие нарушается. Для решения этой проблемы виды адаптируются посредством силы гибридов, когда посторонние гены попадают в генофонд. Мы должны так же помнить, что близкородственное скрещивание не обязательно обозначает, что вредоносные гены будут полностью уничтожены. Гомозиготность повышается, и вредоносные рецессивные аллели вполне могут быть гомозиготными. В реальности, единственный случай, когда вредоносные гены исчезают из популяции, это когда эти аллели выбиваются из популяции путём отбора.
Запомните, что есть два очень отличающихся типа гермафродитов. Постоянные гермафродиты способны производить и яйцеклетки и сперматозоиды одновременно, тогда как мужские гаметы прото-гермафродитов созревают и увядают до созревания женских гамет. Как вы можете догадаться, комбинаций, приводящих к развитию двудомности много. Каждая комбинация развивается по собственному сценарию, который может быть переигран или, что более важно, повёрнута вспять. Это может привести к прорыву в создании двудомных сортов из сортов, у которых есть проблемы с гермафродитизмом. Селекционеры, помните об этом и о различных экспериментах по смене пола.
Целью этой главы было рассказать немного об эволюционном канале, или эволюционном развитии двудомности у растений и угнетении при родственном скрещивании. Всё это связано с условиями выращивания. Так как большинство из нас желает растить одинаковые растения, мы ищем стабильные сорта, даже если они подвергаются угнетению по некоторым признакам. Мы так же ищем двудомные растения, потому что растим сенсимилью, а если мы хотим добавить усиливающей генетики, нам стоит пользоваться преимуществами усиления гибридов. В начале главы мы коснулись глубинных аспектов понимания типов полов конопли, как они развились и как ими можно управлять. Мудрым решением было бы держать все растения, на которых проводились эксперименты с полом подальше от основного генофонда. Однако вы вполне можете поэкспериментировать, получится ли вам избавиться от гермафродитизма в популяции гермафродитов. Это очень хорошее направление работы, особенно если речь идёт о высокоурожайных и мощных гермафродитных сортах, с которыми никто не желает работать из-за невозможности производства сенсимильи.