Кратко о зрении и почему слепота - не болезнь
@neuroslutДумаю, я со временем много буду писать о каких-то деталях исследований, касающихся зрительной системы (мышей в основном). Но, вероятно, имеет смысл сделать какое-то маленькое описание упоминаемых процессов и анатомических образований. Кажется, обывателю может быть сложно с тем, что проблема со зрением может наступить на любом этапе получения и обработки мозгом зрительной информации, и поэтому слепота бывает очень разной.
Я буду опираться на эту обзорную статью о зрении у мышей (все картинки из нее же), но упрощу ее до необходимого минимума. Соответственно, нейробиологам тут вряд ли будет интересно, а вот людям, для которых это легкое хобби, может зайти. Попутно упомяну, как ощущается нарушение зрения при поражении того или иного образования на пути зрительной информации.
Короче, как светит мир во мрак полости черепа?
Приблизительно вот так:
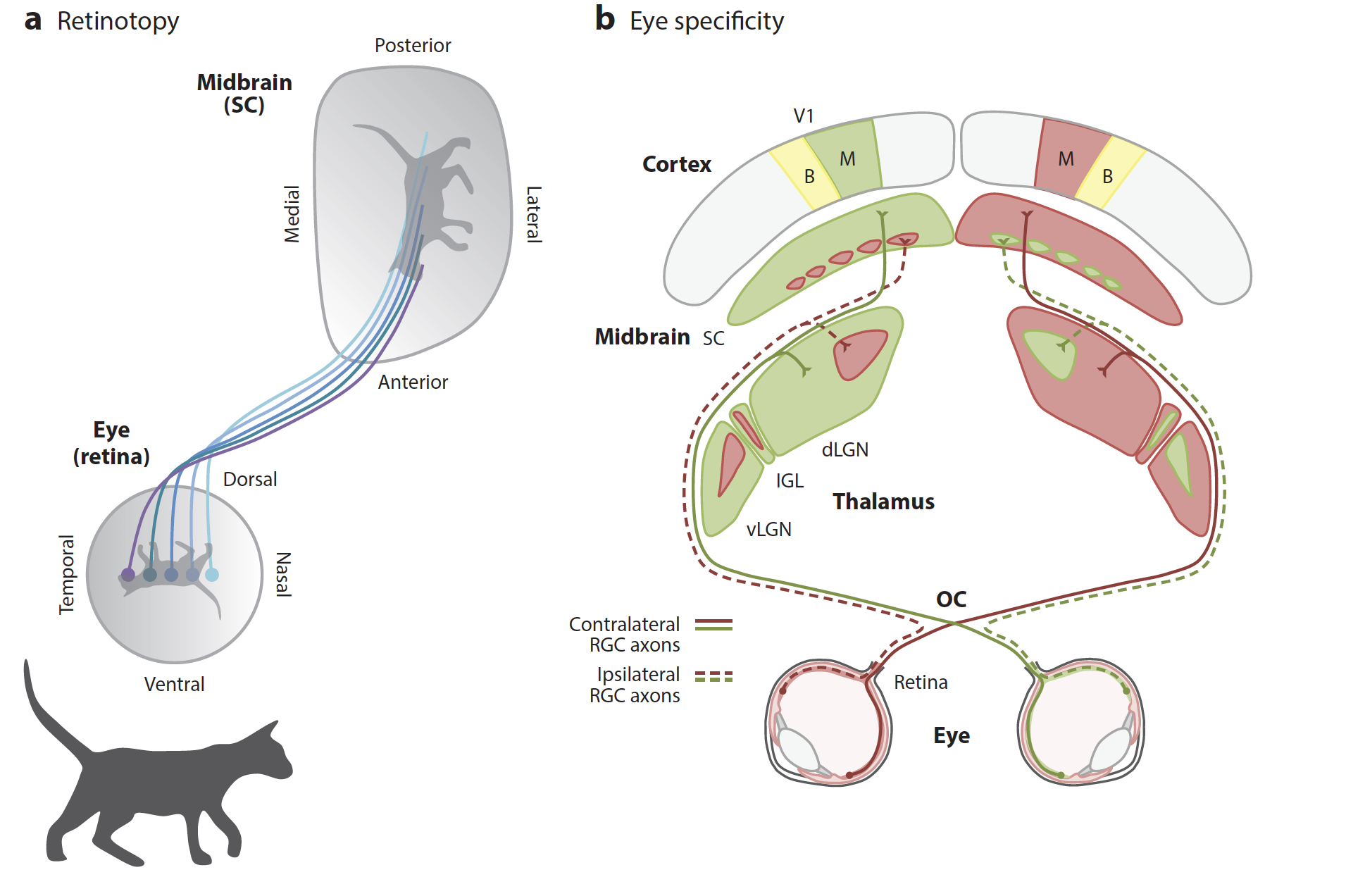
Сокращения: dLGN (dorsal lateral geniculate nucleus) - дорсальное латеральное коленчатое ядро; IGL(intergeniculate nucleus) - межколенчатое ядро; OC(optic chiasm) - зрительный перекрест; RGC(retinal ganglion cell) - ганглионарная клетка сетчатки, та самая, которая "чувствует" падающие на нее фотоны света; SC(superior colliculus) - верхнее двухолмие; V1(primary visual cortex) - первичная зрительная кора; vLGN(ventral lateral geniculate nucleus) - вентральное латеральное коленчатое ядро. M - монокулярное (информация только из 1 глаза), B - бинокулярное.
Зрение на пути в череп
Все начинается в сетчатке, где фоторецепторы (те самые палочки и колбочки, которых у приматов 3 - RGВ, а у мышей только G&UV, т.е. красный цвет мыши не видят), гиперполяризуются от падения на них света соответственной волны, и через несколько клеток-посредников сообщают о свете RGC - первым в этой цепи нервным клеткам, которые реагируют на это деполяризацией - возбуждением, и производят потенциалы действия - разряды, которые по их аксонам дальше идут в мозг, перекрещиваются (но не полностью) и сообщают новость LGN - таламическому центру зрения.
Если повредить кусочек сетчатки 1 глаза, перед взором будет слепое пятно, которое будет очень сложно заметить. Естественно, если больной глаз закрыть, а здоровый оставить открытым, никакого пятна не будет. Также мозг с большой вероятностью "заполнит" дефект картинки тем, что он ожидает там увидеть, а вы этого не будете замечать, пока...
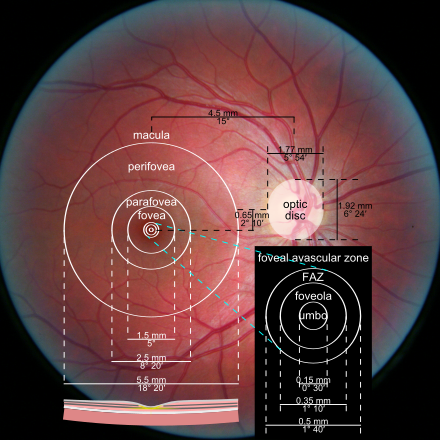
Пока не будет поражена самая важная часть сетчатки - центральная ямка (fovea centralis). Эта маленькая зона загружает информацией более половины первичной зрительной коры, и позволяет нам читать и заниматься другой высокоточной деятельностью.
А теперь прикол: у мышек ее нету. А у ворон есть. А у собак нету. А у варанов есть. Не совсем ясно почему, но из млекопитающих только у высших приматов есть этот хайрез-участок, но у птиц, рептилий, рыб он уже существовал, эволюция вернула нам забытый инструмент. И это здорово мешает делать трансляционные исследования (то есть с планом применить это потом на людях) на других млекопитающих, потому что их философия использования глаз совсем иная, чем у нас.
Теперь посмотрим-ка на человеческое поле зрения:
Фовеальная аваскулярная зона, в которой сосудов нет вообще и самое острое зрение, занимает всего 1.5 градуса поля зрения. Это примерно площадь ногтя большого пальца на вытянутой перед собой руке. Не очень много, правда? Если быть щедрым и говорить о макулярном зрении, то это около 17-20°, но это уже гораздо более широкое определение, потому что макула в целом видит в меньшем разрешении, чем фовеа. Приматы используют саккады - небольшие движения глаз, похожие на мазки краской у художника, чтобы уловить все детали интересующего нас предмета. Нарушения зрения, которые связаны с невозможностью исполнять саккады или в принципе содружественно двигать глазами, я тут не трогаю. Скажу только, что саккадируем и сознательно двигаем глазами мы под контролем отдельного участка мозга, который вообще расположен в лобной доле - frontal eye fields. Да, это далековато от зрительной коры в затылке, но зато близко к нашему исполнительному директору - префронтальной коре.
Слепота от повреждения сетчатки бывает от массы причин: отслоение сетчатки из-за травмы, меланома сетчатки, поражение сосудов (например, из-за диабета), а также десятки других причин - например, если вы повидали слишком много дерьма (то есть ультрафиолета) за свою жизнь. Но если повреждена фовеальная часть сетчатки, саккады не дают острого зрения, и это будет очень портить жизнь, в то время как повреждение более обширного участка на периферии может быть вообще незаметно пациенту.
Слепота от повреждения зрительного нерва - пучка отростков нервных клеток из сетчатки в мозг - редко бывает по типу "выбитых пикселей", чаще она полная. Зрительный нерв очень чувствителен ко всему. Например, один из моих пациентов ослеп после облучения по поводу опухоли центральной нервной системы. Но в целом это возможно, например, при неполном перерезании. Но крайне маловероятно.
Зрение внутри черепа
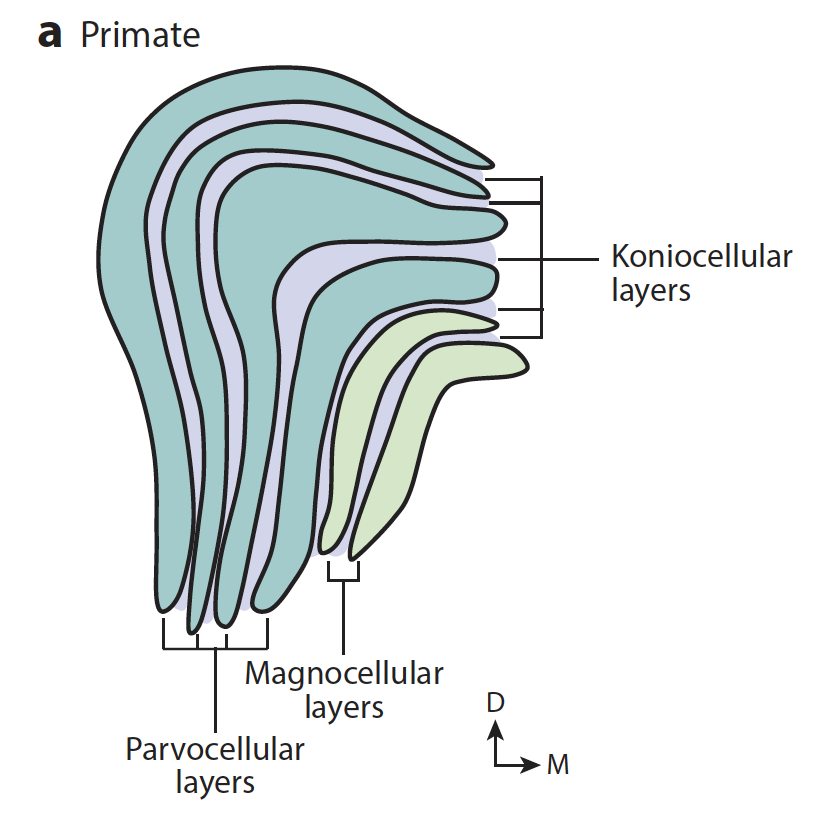
LGN мышей раньше считалась таким простым реле информации из глаза дальше в мозг, однородным по строению, в то время как у приматов (и некоторых других высокоразвитых млекопитающих) это образование слоистое, как торт Наполеон, и разные слои делают разные вещи: одни (М-клеточные) проводят быструю монохромную(ч/б) контрастную информацию, другие (П-клеточные) проводят медленную цветную информацию в высоком разрешении. М и П сами по себе связаны со специфическими подтипами RGC клетчатки, но и в LGN это разные на вид клетки.
Но теперь у нас есть новые возможности отследить различия в форме и генетических маркерах RGC мышей, и оказалось, что у них тоже есть разные такие пути внутри дорсального LGN:
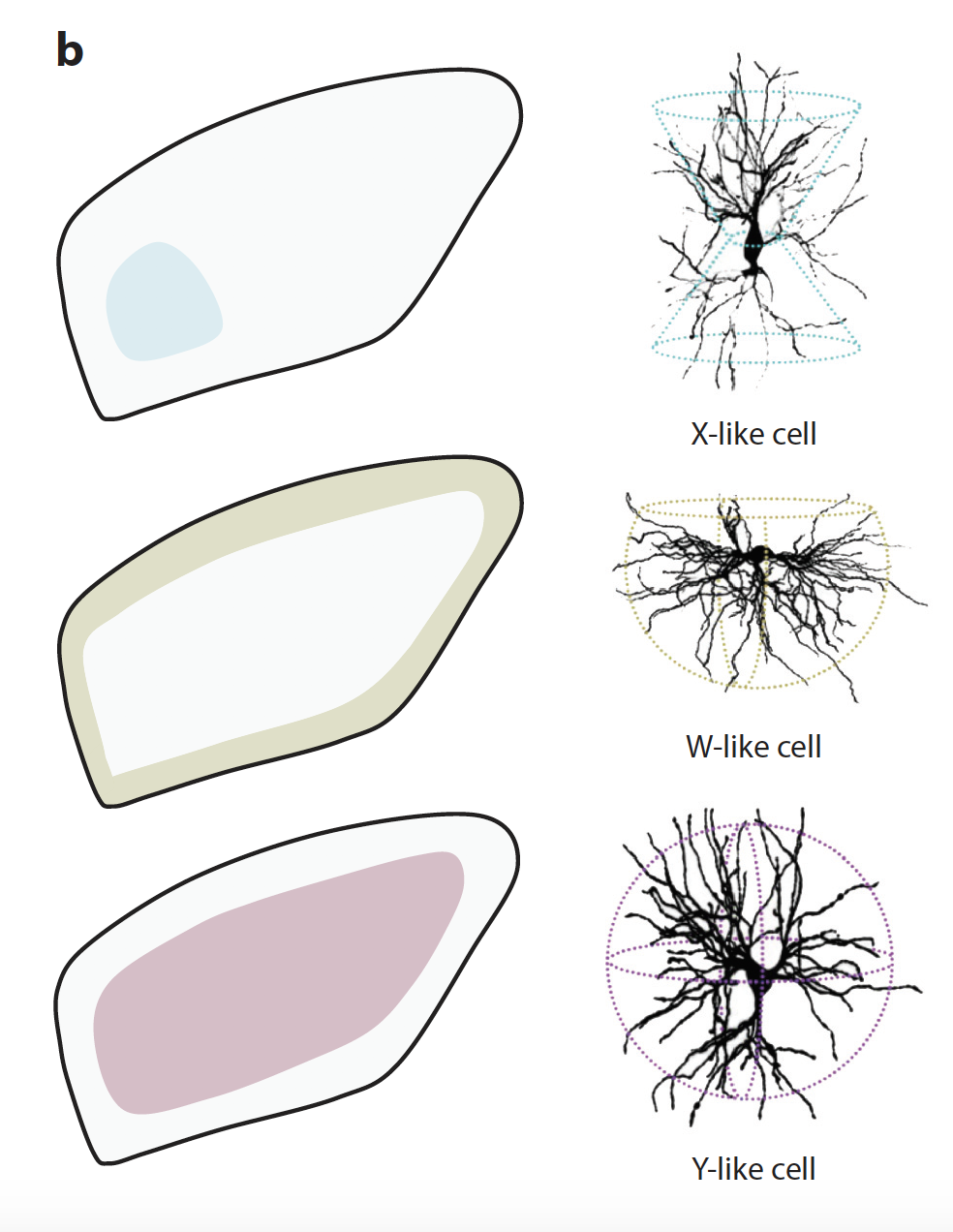
Тем не менее, грызунам далеко до слоистости LGN приматов и хищников (у собак и кошек оно тоже слоистое, то есть разделено на отдельные каналы передачи информации).
Так мы добрались до второго нейрона зрительного пути и дальше, и тут проще показать:
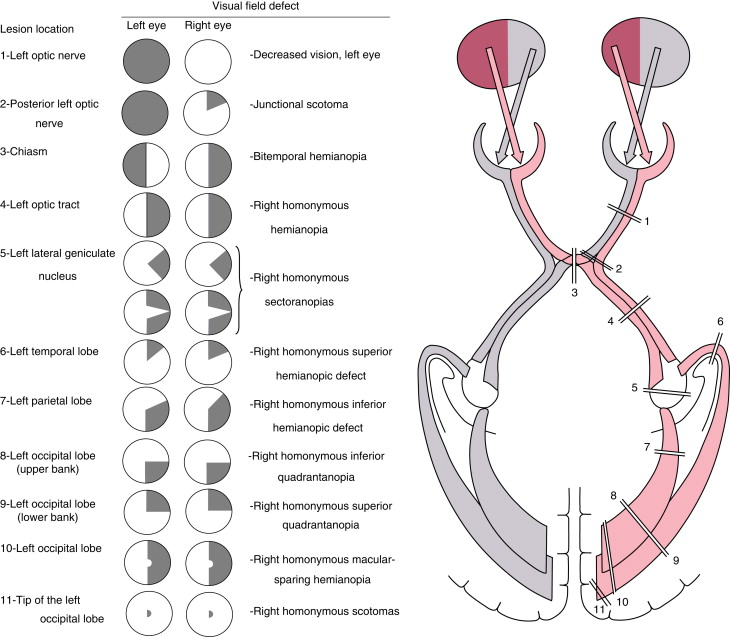
Важно, что у человека зрительные нервы перекрещиваются и информация с обоих глаз распределяется в полушария относительно hemifield - того, какая это половина условно видимой глазу левой/правой полусферы, и всё латеральное полуполе (внутренняя половина обеих сетчаток) направляется к противоположному по стороне (контралатеральному) полушарию к зрительной коре. На самом деле именно детальное описание того, какая часть поля зрения "ослепла", помогает довольно точно предугадать место поражения в зрительной системе. Но возятся с этим не так часто, потому что прокатать человека на МРТ быстрее и доказательнее, а это - лишь подсказка, где искать (например, при выпадании обоих внешних hemifields мы будем делать прицельный снимок гипофиза, потому что его опухоли чаще всего давят на перекрест зрительных нервов).
Так, мы видим принцип ретинотопии: то, какая часть элемента зрительной системы повреждена (или стимулирется), будет определять, какая часть поля зрения от этого выпадет (или почувствует стимуляцию). Мы также видим, что ретинотопия - свойство не только первичной зрительной коры, но и всех структур начиная с сетчатки. Это свойство общее для всех животных со сложной зрительной системой, которых мы знаем (головоногих не считаем).
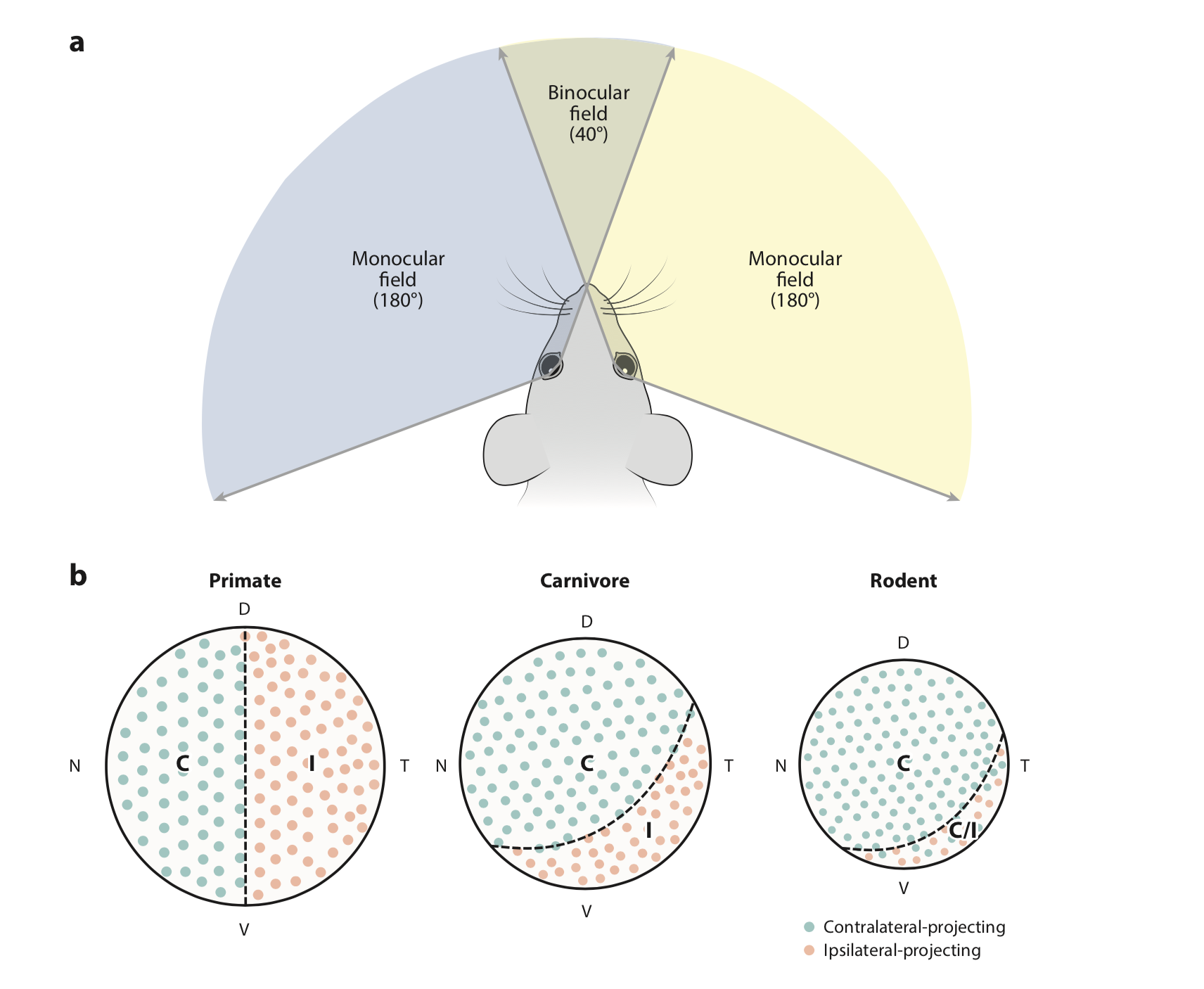
У человека и хищников каждый участок сетчатки связан с конкретным полушарием в зависимости от hemifield, но у мышей та часть поля зрения, которая видна обоим глазам, отправляется в оба полушария. Это, как кажется, сложнее, чем у приматов, ведь части нейронов приходится совмещать сигналы из обоих глаз, а части - нет. А у приматов вся зрительная кора совмещает сигналы из только левых или только правых половин обеих сетчаток, и это реализуемо уже на этапе LGN, поэтому оно слоистее, но в некотором смысле однороднее. Принцип ретинотопии сохраняется независимо от того, бинокулярное ли зрение у этого вида.
Зрение внутри первичной зрительной коры
Я зачем-то упоминала о разных слоях LGN, которые делают разные вещи: М-клеточные (ч/б хай-рез), П-клеточные (цветное, медленное), и ещё у приматов есть К-клеточный, он как кремовая прослойка между М- и П- и мы не до конца понимаем, что он делает.
У приматов П-клетки (парвос - маленький, потому что эти клетки в микроскопе маленькие) LGN посылают аксоны в первичную зрительную кору не равномерно, а в виде цилиндрообразных скоплений, которые называются блобами (blobs), потому что похожи на более темные пятна, если покрасить кору так, чтобы было видно активность энзима цитохром оксидазы.
У приматов в блоб направляются аксоны П-слоя, а в интерблобы - М-слоя (магноцеллюлярного, потому что эти клетки крупнее), хотя Callaway почему-то думал иначе. Условно говоря, блобом мы анализируем цвет и форму, но темпоральное разрешение страдает, поэтому эти участки можно назвать медленными. Если разрушить П-клеточный участок LGN, у пациента будут сложности с восприятием цвета и мелких деталей, и блобы через некоторое время исчезнут или будут неопределимыми на микроскопии коры. Если разрушить М-клетки, то ухудшится контрастность изображения и потеряется темпоральное разрешение, поэтому мы хуже будем распознавать границы предметов, а также движения в поле зрения. То есть, условно говоря, как хищнику и как добыче, коими мы являлись, нам вроде как важнее М-клеточный путь, но чтобы отличить спелый фрукт от неспелого или заприметить замершего тигра в засаде, нам нужен П-путь. Они на самом деле работают сообща, хотя считается, что М-путь сообщает нам, куда саккадировать, а также дает ощущение глубины как бинокулярному виду.
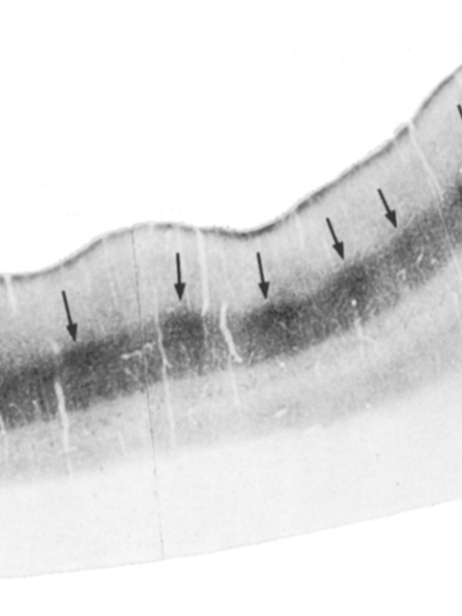
У грызунов блобов нет, хотя есть очевидно разные виды клеток в сетчатке и в LGN. Грызуны решили, что им проще анализировать картинку тотально, и поэтому "блобность" у них по типу перца с солью, распределенная равномерно, а не в виде пятен. И при том, что это вроде как отличие, мешающее делать исследования, которые проще перенести на приматов, это на самом деле может быть небольшим примуществом: если мы делаем нейропротез на зрительной коре, то разница в десятки микрометров может случайно сделать его более блобно-расположенным у одного животного и более интерблобно - у другого, и это может добавить вариативность результатам, хотя вроде как ныне существующие электростимулирующие нейроинтерфейсы на практике такой проблемы не имеют. Но я верю, что когда-то мы достигнем такой точности, что реально придётся задуматься над блобностью.
Таким образом, зрительная система достаточно многогранна, чтобы вместо "слепоты" и даже "корковой/окулярной слепоты" придумать под 20 названий разных "слепот", которые даже не отображают патофизиологию, а только топику повреждения. И конечно же легальное понимание слепоты скорее применимо к окулярной, "обычной" слепоте, чем к центральным ее разновидностям. Животные модели полны нюансов, разных для разных видов, и даже учет всех этих особенностей не гарантирует трансляционного потенциала таких исследований, то есть их применимость к людям очень ограничена. Короче, мне жаль, что в 21 веке так сложно делать нейропротезирование на людях как единственной достоверной модели человеческого зрения, но радостно, что мы знаем так много о строении зрения других животных.