Исследование показало как псилоцибин вызывает столь важные изменения в работе мозга, что перезагружает его
MUSHROOM GENETICS
miceliar
💡 Отказ от ответственности
В твоем мозге нет кнопки сброса. Но чем больше ученые узнают о волшебных грибах, тем больше убеждаются, что они настолько близки к кнопке перезагрузки, насколько это возможно.
Учёные создали визуализацию работы мозга под псилоцибином, и представили рабочую модель на основе которой становится понятно, если "кнопка сброса настроек" для мозга и существует - эта кнопка псилоцибин.
Построенная модель человеческого мозга под псилоцибином проливает свет на то, как галлюциногенные грибы позволяют нашему мозгу получить доступ к неиспользованному потенциалу. Эта модель показывает, что под влиянием псилоцибина мозг создает петлю обратной связи активности нейронов и высвобождения нейротрансмиттеров (химических мессенджеров, которые нейроны используют для связи). Модель так же демонстрирует что псилоцибин может позволить мозгу ПОДКЛЮЧИТЬСЯ К ИНТЕРАКТИВНЫМ НЕДОСТУПНЫМ СОСТОЯНИЯМ, включая «дестабилизацию» отдельных мозговых сетей и создание более «глобальной» сети по всему мозгу.
«Использование этой модели будет иметь решающее значение для понимания того, как псилоцибин может сбалансировать нейропсихиатрические расстройства, такие как резистентная к лечению депрессия и зависимость», — говорит Кринглебах руководитель исследования.

В течение обычного дня в человеческом мозге нейроны постоянно возбуждаются, а нейротрансмиттеры путешествуют по проторенным дорожкам через мозг, что-то вроде автомобилей на автостраде. На галлюциногенных грибах эти сети «дестабилизируются» - объяснил он.
Исследования показали, что новые сети появляются одновременно. Это как если бы автомобили на автостраде получили полную свободу действий, чтобы отклониться от шоссе и свернуть на проселочные дороги в направлении новых пунктов назначения.
У нас есть только фиксированное количество "оборудования" в мозге, но мы запускаем очень сложное программное обеспечение, которое производит сны, сознание и — если кто-то принимает вещество, такое как псилоцибин — «прорывные переживания».
Если волшебные грибы что-то и демонстрируют, так это то, что мозг может научиться использовать свое фиксированное оборудование совершенно по-разному, если задействованы правильные ингредиенты. Хитрость заключается в том, чтобы выяснить, какие инструменты нужны мозгу для запуска различных типов программного обеспечения на этом оборудовании.
Текст ниже прямой перевод результатов исследования, он пестрит терминами и довольно сложен для понимая, если Вам интересно понять саму структуру исследования, можете прочитать, если же хотите прочитать выводы из исследования перейдите по ссылке, она перебросит Вас в конец документа к выводам, но так Вы пропустите все картинки. (ссылка).
Исследование было опубликовано в Proceedings of the National Academy of Sciences. Переведено специально для MUSHROOM GENETICS.
Создать столь полную картину работы мозга и обрисовать двунаправленную моделирующую динамическую взаимную связь между нейронной и нейротрансмиттерной системами удалось благодаря объединению данных мультимодальной нейровизуализации (диффузионная магнитно-резонансная томография [dMRI], функциональная магнитно-резонансная томография [fMRI] и позитронно-электронная томография [PET].
Основная цель модели взаимосвязанных нейронов и нейротрансмиттеров всего мозга (показана на рис. 1) состоит в том, чтобы исследовать тесные взаимодействия между этими двумя разными, но взаимно связанными динамическими системами всего мозга (рис. 1А), которые подобраны (рис. 1B) к данным эмпирической нейровизуализации BOLD (рис. 1C).
Другими словами, модели только нейронов, которые соответствуют данным нейровизуализации с использованием динамики локальных узлов, ограниченной анатомической структурной связностью, теперь описываются сбалансированной динамической моделью среднего поля, которая последовательно выражает временную эволюцию ансамблевой активности различных популяций нейронов, формирующих сеть шипов. Спонтанная активность каждой отдельной области мозга описывается сетью, состоящей из двух пулов возбуждающих и тормозных нейронов (рис. 1D). Система нейротрансмиттеров, с другой стороны, использует отдельный набор дифференциальных уравнений, описывающих динамику уровня концентрации нейромедиатора, задаваемую хорошо известной динамикой высвобождения и обратного захвата Михаэлиса-Ментен. На этом этапе учёные связывают нейротрансмиттерные и нейронные динамические системы с помощью анатомической связи между ядром шва и остальной частью мозга, оцененной с использованием dMRI проекта Human Connectome Project (HCP). Явная связь между нейротрансмиттером и нейронной системой показана на рис. 1F . Как видно, токи нейротрансмиттеров применяются к возбуждающим и тормозным пулам нейронов каждой области с использованием параметров эффективности/проводимости (WE и WI соответственно). В каждой области потоки нейротрансмиттеров также масштабируются по плотности рецепторов каждой области (измеренной in vivo с помощью ПЭТ). Обратная связь между нейронной и нейротрансмиттерной системой определяется путем вставки в уравнение Михаэлиса-Ментен высвобождения и обратного захвата скорости возбуждения популяции нейронов в области мозга-источнике нейротрансмиттера, распространяемого из ядра шва.
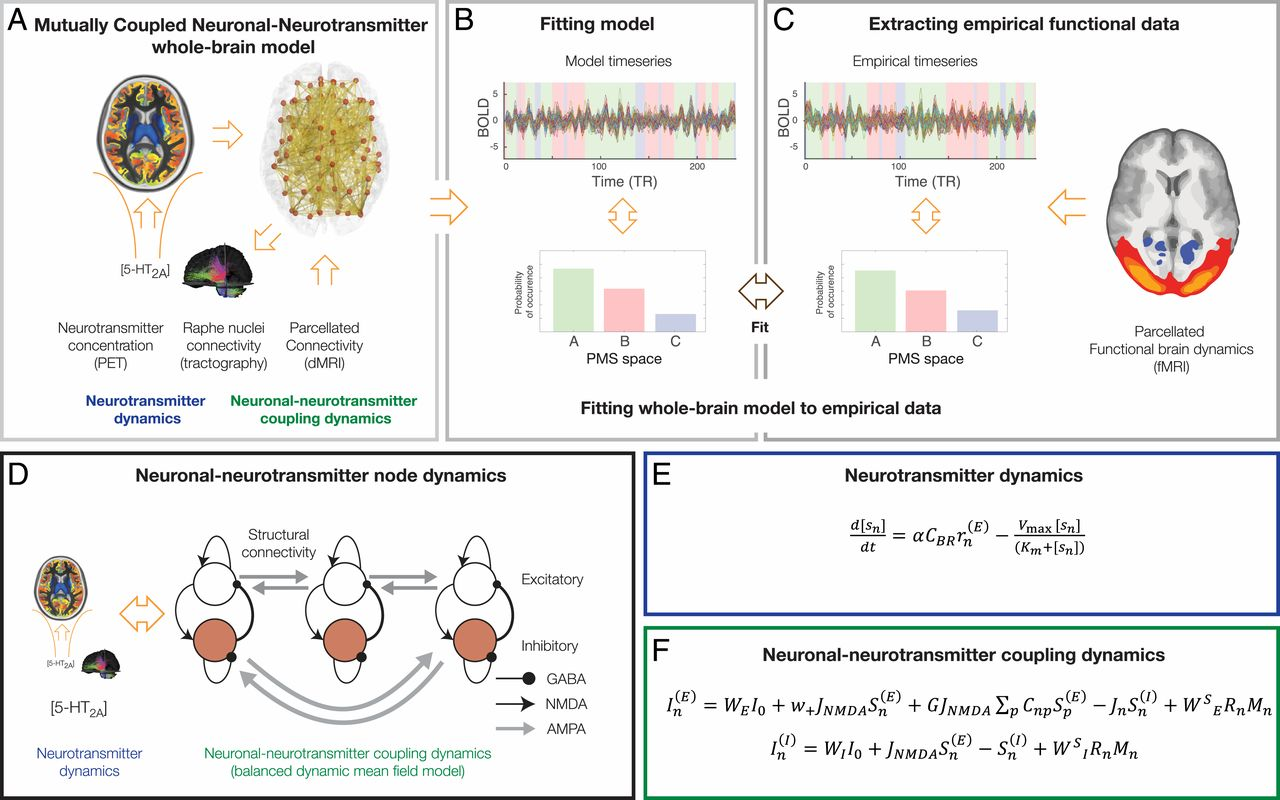
(A) Мы изучили взаимную связь двух различных взаимосвязанных динамических систем всего мозга (нейронной и нейротрансмиттерной). (B) Эта система соответствует эмпирическим данным нейровизуализации, которые описываются пространством вероятностных метастабильных подсостояний (PMS) (C), которое извлекается из эмпирических данных BOLD. (D) Мы достигаем этого, добавляя связанную систему нейротрансмиттеров (синяя) для модуляции и взаимодействия с нейронной системой (зеленая), которая была смоделирована с использованием сбалансированной динамической модели среднего поля, которая последовательно выражает временную эволюцию ансамблевой активности различных нейронных популяций, формирующие спайковую сеть. (E) Система нейротрансмиттеров (синяя) моделируется набором дифференциальных уравнений, описывающих динамику уровня концентрации нейротрансмиттера, которая дается известной динамикой высвобождения и обратного захвата Михаэлиса-Ментен. (F) Динамика связи нейронов (зеленый) моделируется другим набором дифференциальных уравнений, описывающих спонтанную активность каждой отдельной области мозга, состоящей из двух пулов возбуждающих и тормозных нейронов. Мы связываем нейротрансмиттерные и нейронные динамические системы через анатомическую связь между ядром шва и остальной частью мозга, оцененную с помощью dMRI от HCP. Явная связь между нейротрансмиттером и нейронной системой представлена в уравнениях.
Как можно ясно видеть, токи нейротрансмиттеров применяются к возбуждающим и тормозным пулам нейронов каждой области с использованием параметров эффективности/проводимости (WE и WI соответственно). В каждой области потоки нейротрансмиттеров также масштабируются по плотности рецепторов каждой области (измеренной in vivo с помощью ПЭТ). Обратная связь между нейронной и нейротрансмиттерной системой определяется путем вставки в уравнение Михаэлиса-Ментен высвобождения и обратного захвата скорости возбуждения популяции нейронов в области мозга-источнике нейротрансмиттера, распространяемого из ядра шва.
В качестве демонстрации силы этой общей схемы исследователи показывают, как ее можно использовать для объяснения конкретного случая модуляции серотониновой системы псилоцибином путем анализа данных нейровизуализации группы здоровых участников, которым вводили псилоцибин внутривенно.
Это позволило изучить соответствующую роль связи между системами нейронов и нейротрансмиттеров, чтобы понять специфичность плотности рецепторов 5-HT2A [составлено с помощью ПЭТ in vivo]. В частности, основным источником нейротрансмиссии серотонина является ядро шва, которое необходимо для правильной связи между обеими динамическими системами. Для этого использовали структурную связность из этой области, оцененную с помощью трактографии dMRI .
Учёным удалось подогнать взаимно связанную модель всего мозга, используя схему описания состояния мозга как ансамбля или вероятностного «облака» в заданном пространстве состояний. Это облако, конечно, не сгруппировано в дискретные состояния, но было показано, что кластеризация, тем не менее, может быть полезна для обеспечения так называемых «метастабильных подсостояний», которые могут значительно различать состояния мозга.
Состояние мозга определяется набором метастабильных подсостояний, то есть изменяющихся во времени псевдосостояний, возникающих в результате процесса кластеризации.
Исследователи извлекли это пространство вероятностных метастабильных подсостояний (PMS) для эмпирических данных о псилоцибине (плацебо и активное состояние) с использованием метода анализа динамики ведущих собственных векторов (LEiDA) (схематически представленного на рис. 2).
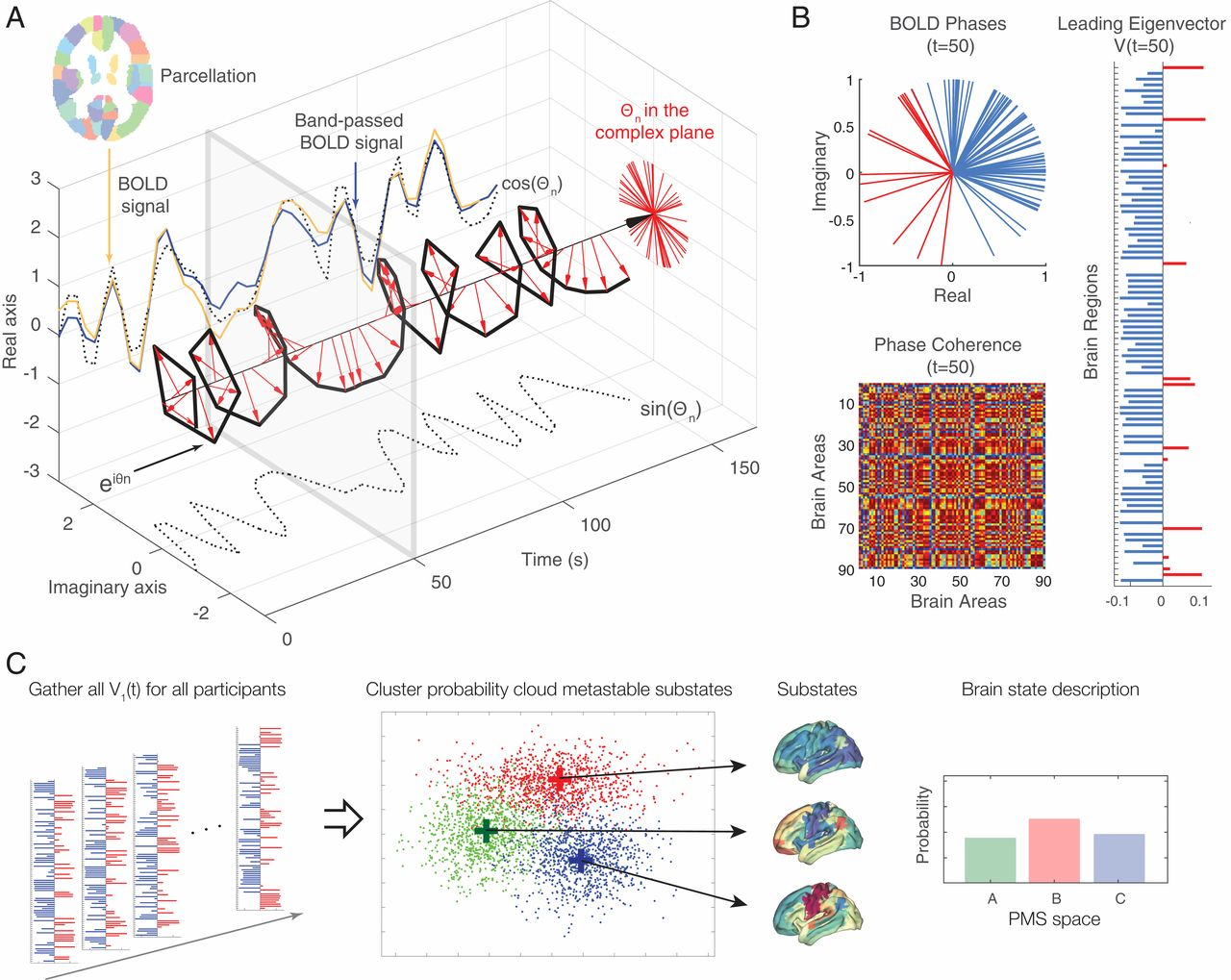
Вкратце, мы определяем динамическую матрицу FC (dFC) с временным разрешением, используя BOLD связность фазовой когерентности. Чтобы уменьшить размерность задачи, мы вычисляем соответствующий ведущий собственный вектор с временным разрешением, который фиксирует доминирующий шаблон связности dFC (t) в момент времени t. Затем мы обнаруживаем дискретное количество метастабильных подсостояний путем кластеризации dFC(t) по временным точкам и субъектам. Полученные k кластерных центроидов определяют пространство PMS, для которого мы вычисляем вероятности, время жизни и вероятность перехода между ними как для плацебо, так и для активных условий псилоцибина. Плацебо и активные условия псилоцибина можно существенно отличить по трем подсостояниям.
На рис. 3А показаны три различных подсостояния методологии ПМС с соответствующими вероятностями и вероятностями перехода между ними для плацебо и активных состояний псилоцибина. Как видно на подграфиках, два подсостояния (1 и 3) значительно различаются между двумя условиями (P < 10–4) с точки зрения вероятности, а подсостояние 3 значительно отличается по продолжительности жизни (P < 10–2). Это демонстрирует, что кластерный подход действительно полезен для различения состояний мозга.
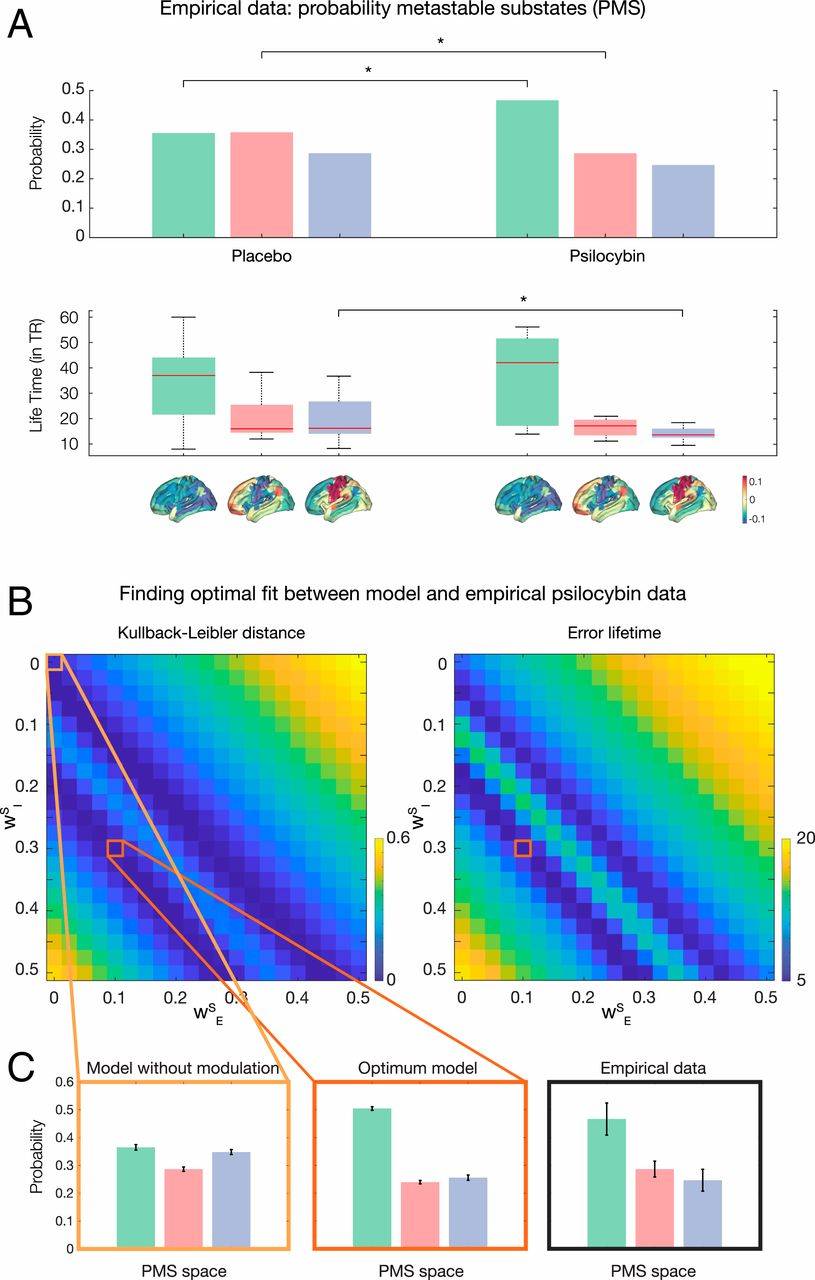
Сначала учёные приспособили модель всего мозга к пространству PMS плацебо-состояния псилоцибина, используя только нейронную систему (и, таким образом, без связи с нейротрансмиттерной системой). Они сделали это, установив минимум симметричного расстояния Кульбака-Лейблера (KLD) между эмпирическим плацебо-состоянием PMS и смоделированным PMS. Это дало глобальный параметр связи нейронов G = 1,6. В равной степени мы измерили энтропийное расстояние Маркова (МЭ). Оптимальными оказались значения KLD = 0,002 и ME = 0,05.
Во-вторых, исследователи приступили к изучению роли динамической системы нейротрансмиттеров, объединив обе системы в модель. Это позволило предсказать изменения динамики мозга под действием псилоцибина. В частности, во взаимно связанной модели всего мозга использовались те же центроиды, что и в эмпирических данных нейровизуализации, и двумерная (2D) матрица параметров связи WSE и WSI для создания смоделированного PMS (рис. 3B). Мы искали эту матрицу для оптимальной подгонки (глобальный минимум) к активным условиям данных о псилоцибине, используя оптимальный симметричный KLD между эмпирическим активным состоянием псилоцибина PMS и смоделированным PMS. Тем не менее, из глубокого диагонального выступа ясно, что существует инвариантное соотношение между WSE и WSI, которое дает эквивалентные значительно отличающиеся результаты в отношении WSE=0 и WSI=0, что является релевантным сравнением. На рис. 3B показаны матрицы KLD и время жизни ошибки подсостояний в эмпирическом псилоцибин-активном состоянии PMS и смоделированном PMS в зависимости от параметров связи WSE и WSI. Оптимальное описание состояния псилоцибиновой активности находится при минимуме KLD при WSE=0,3 и WSI=0,1, а полное отключение нейромедиаторной системы – при WSE=0 и WSI=0. На рис. 3C показаны пространства PMS для несвязанной системы (WSE = 0 и WSI = 0, крайняя левая панель), оптимальное соответствие модели (WSE = 0,3 и WSI = 0,1, средняя панель) и эмпирические данные (крайняя правая панель).
При таком оптимальном соответствии взаимно связанная модель всего мозга позволяет получить дальнейшее представление о лежащей в основе динамике нейротрансмиссии, связанной с псилоцибином (в данном случае с серотонином). На рис. 4А показана значительная разница между оптимальной посадкой и несвязанной системой при КЛД (P < 10–6). Это ясно демонстрирует важность сопряжения нейронной и нейротрансмиттерной систем. Рис. 4B дополнительно анализирует это открытие, показывая значительную разницу между оптимальной подгонкой и оптимальной подгонкой, но при условии чито в ней заморозили динамику обратной связи от системы нейротрансмиттеров к нейронной системе (P < 10-6). Это было сделано путем разрешения связи до тех пор, пока не будет достигнуто устойчивое состояние, а затем выбрано среднее значение переменных нейротрансмиттеров, заморажено и отменена динамика обратной связи. При таком разобщении исследователи рассматривают более простую связанную систему, в которой динамика нейронов сохраняется, но не может влиять на замороженную динамику нейротрансмиттеров.
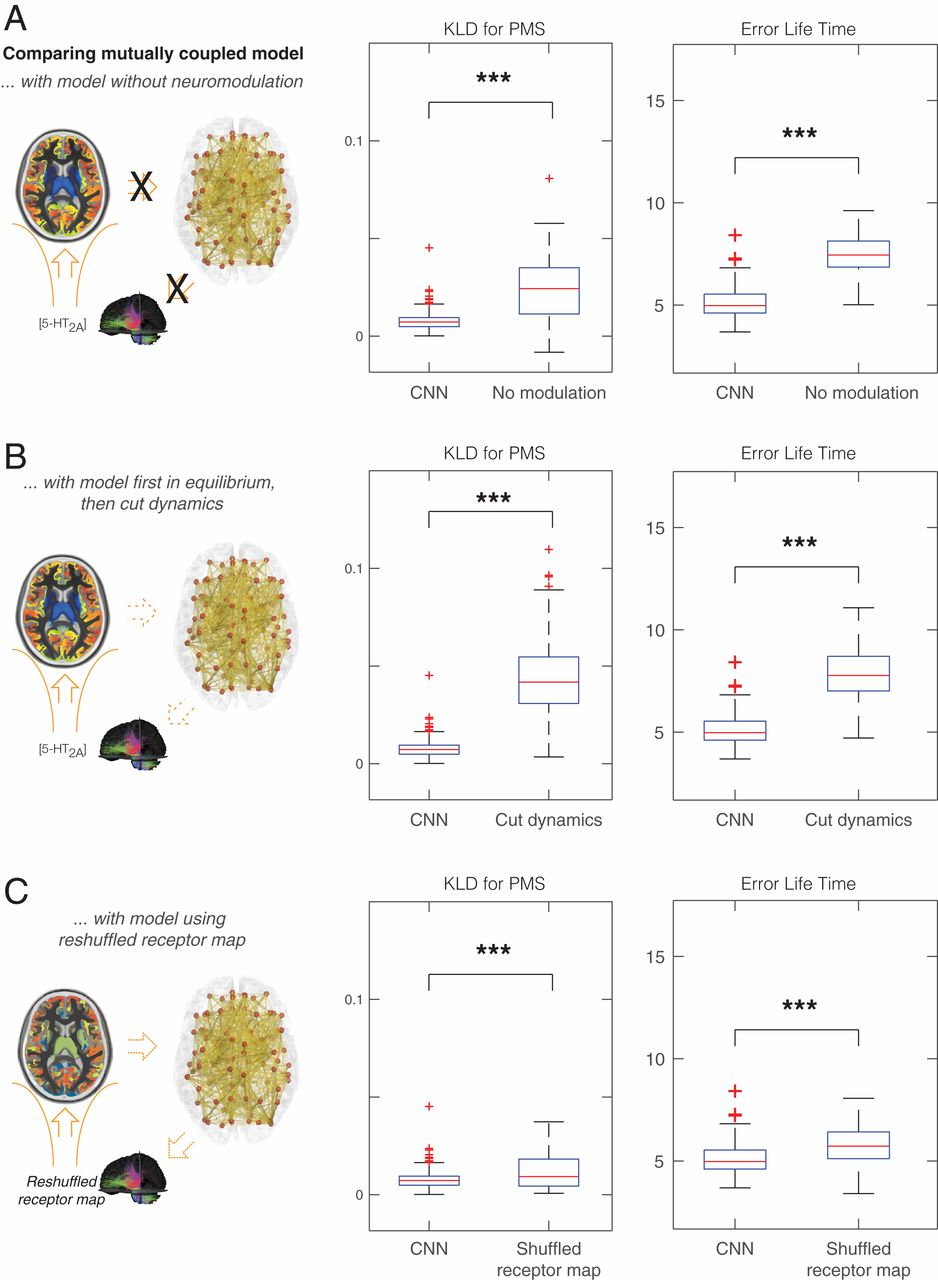
На рис. 4C показана значительная разница между использованием эмпирических плотностей рецепторов 5-HT2A по областям при оптимальной подгонке по сравнению с результатами случайной перетасовки плотностей рецепторов (P < 10–4).
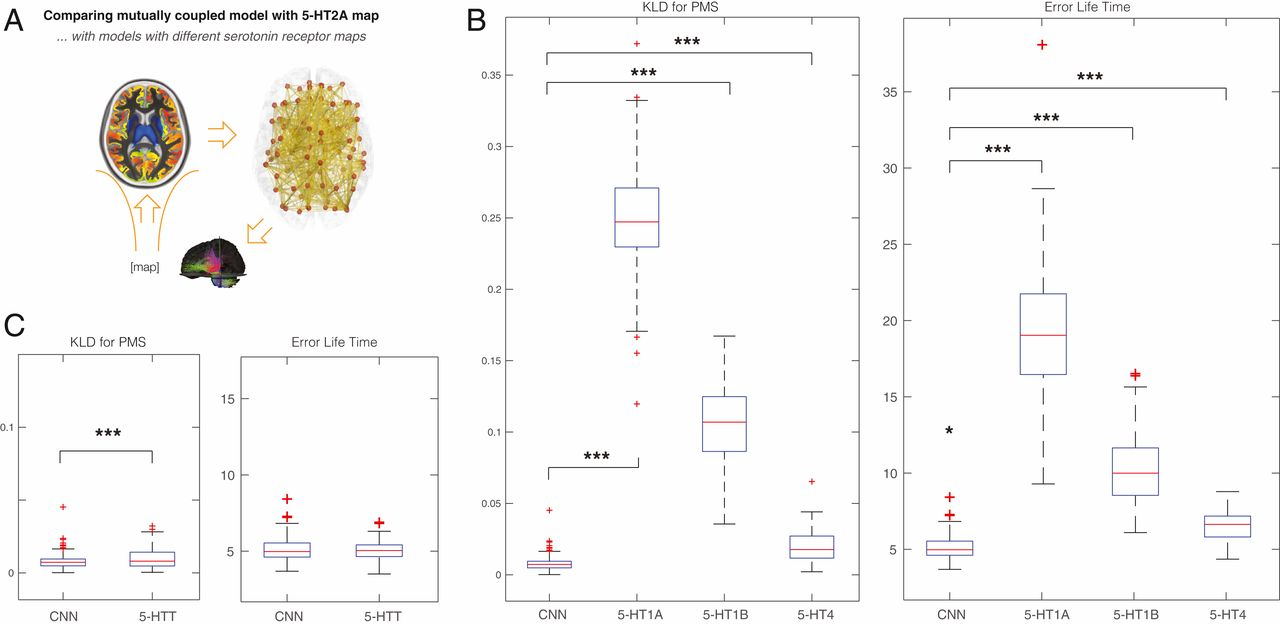
Точно так же, это показано на рис. 5А, где исследователи также проверили специфичность карт связывания рецепторов, сравнив (при оптимальной подгонке) рецептор 5-HT2A с другими рецепторами серотонина, а именно 5-HT1A, 5-HT1B и 5-HT1B. HT4, который показал значительные различия между 5-HT2A и 5-HT1A (P <10-6), 5-HT1B (P <10-6) и 5-HT4 (P <10-6). Это убедительно подтверждает основную роль этого рецептора в эффектах псилоцибина.
Наконец, на рис. 5B мы показываем сравнение с 5-HTT (траспортёр серотонина), которое также значимо (P <0,02). 5-НТТ не является рецептором, но было показано, что он играет важную роль в лечении депрессии.
Выводы исследователей
Здесь мы показали, как динамическая взаимосвязанная модель всего мозга может решить главную проблему нейробиологии, которая состоит в объяснении парадоксальной гибкости функций человеческого мозга, несмотря на зависимость от фиксированного анатомического коннектома.
Одним из наиболее важных элементов этой гибкости является способ модуляции фиксированного коннектома нейромодуляторами для избирательного изменения баланса возбуждения и торможения отдельных областей мозга.
Здесь мы смоделировали динамическую взаимную связь между системами нейронов и нейромодуляторов на уровне всего мозга. В частности, мы внедрили взаимно связанную динамическую систему, которая связывает на уровне всего мозга нейронную систему, которая была смоделирована с использованием сбалансированной динамической модели среднего поля, и систему нейромодуляторов, описывающую динамику уровней концентрации нейротрансмиттеров. (измерено in vivo с помощью ПЭТ) и смоделировано с помощью хорошо известной динамики высвобождения и обратного захвата Михаэлиса-Ментен.
Здесь, в качестве доказательства принципа, мы рассматриваем влияние псилоцибина на систему серотонина и поэтому используем анатомическую связь между ядром шва и остальной частью мозга, чтобы соединить две системы.
В целом результаты показывают, что взаимодействие между этими динамическими системами является фундаментальным для объяснения эмпирических данных. Другими словами, динамическое взаимодействие между нейронной и нейромодуляторной системами на уровне всего мозга важно для полного объяснения функциональной модуляции мозговой активности псилоцибином, мощным психоделическим препаратом, действующим на серотониновую систему.
Это особенно важно, учитывая продемонстрированную способность псилоцибина восстанавливать баланс человеческого мозга при резистентной к лечению депрессии. Результаты свидетельствуют о том, что интеграция дМРТ (анатомия), фМРТ (функциональная активность нейронов) и ПЭТ (нейротрансмиттерная система) на уровне всего мозга необходима для правильного прогнозирования динамики мозга в результате взаимной связи между двумя динамическими системами. Это расширяет существующие обширные экспериментальные и теоретические исследования локальных эффектов нейротрансмиттеров.
В частности, результаты дают представление об основной динамике нейротрансмиссии, связанной с псилоцибином. Что касается динамики, мы сначала отделили нейромодуляторы от нейронных систем и обнаружили, что это привело к очень значительному нарушению соответствия эмпирическим данным (рис. 4А). Затем мы провели дальнейшее моделирование, чтобы исследовать роль конкретных частей динамической связи. Полная взаимно связанная модель всего мозга работала до тех пор, пока не было достигнуто устойчивое состояние, после чего мы заморозили динамику обратной связи от нейромодуляторов к нейронной системе, удалив связь через ядро шва. Это снова привело к очень значительному нарушению соответствия эмпирическим данным (рис. 4B).
В дальнейших сериях экспериментов, предназначенных для изучения важности распределения рецепторов, мы изменили распределение плотности региональных рецепторов путем 1) случайного перетасовки 5-HT2A (рис. 4C) и 2) замены их на другие известные рецепторы серотонина. быть гораздо менее чувствительными к псилоцибину, а именно 5-HT1A, 5-HT1B и 5-HT4. Опять же, это привело к значительному нарушению способности объяснить эмпирические данные (рис. 5В). Этот результат подтверждает решающую причинную роль точного анатомического расположения и плотности 5-HT2A.
Интересно, что, как упоминалось выше, было показано, что псилоцибин играет роль в восстановлении баланса человеческого мозга при резистентной к лечению депрессии. Считается, что терапевтическое действие псилоцибина при депрессии зависит от активации рецепторов серотонина 2А, тем самым инициируя многоуровневую пластичность. Это отличается от селективных ингибиторов обратного захвата серотонина, которые являются наиболее часто назначаемым классом антидепрессантов и терапевтическое действие которых, как считается, начинается с блокады обратного захвата в 5-НТТ; псилоцибин не имеет заметного сродства или действия на 5-HTT. Мы были заинтересованы в дальнейшем изучении этого, заменив плотности рецепторов на плотности 5-HTT. Мы обнаружили, что соответствие с 5-HTT было значительно хуже, чем с 5-HT2A (рис. 5C), и подтверждает потенциал псилоцибина в восстановлении баланса функций мозга при резистентной к лечению депрессии.
В более широком смысле, взаимосвязанная модель всего мозга и полученные результаты проливают важный свет на наше понимание функций человеческого мозга. За последнее десятилетие открытие моделирования всего мозга привело к новым важным открытиям для объяснения принципов работы человеческого мозга на основе данных нейровизуализации человека. Первоначальным прорывом стало признание того, что анатомическая связь форм человеческого мозга функционирует на уровне всего мозга. Однако также относительно быстро стало ясно, что это было побочным продуктом тесной связи между анатомией и функцией, учитывая, что можно описать статическую спонтанную активность.
Другие интересные статьи:
🗡The Guardian: Исследование показало, что грибы — самый безопасный рекреационный наркотик
🔞 Секс под грибами. Многие думали, немногие решились.
🧠Психоделические грибы от депрессии: это было «тем, что изменило всё».
Приобрести мицелий Golden Teacher, Pink Buffalo, White Teacher, Penis Envy Uncut, Long Monkey Penis и многих других сортов можно в боте.

👉 https://t.me/joinchat/AAAAAFOVHll23fUPh1flnQ
ПОЖАЛУЙСТА, указывайте ссылку на наш канал при репосте, а лучше просто пересылайте наш пост к себе, мы вкладываем в написание статей силы и время, а Вы получите плюсик в карму если поможете нам.