ЭТАПЫ И ОЧАГИ ОБРАЗОВАНИЯ ЧЕЛОВЕЧЕСКИХ РАС
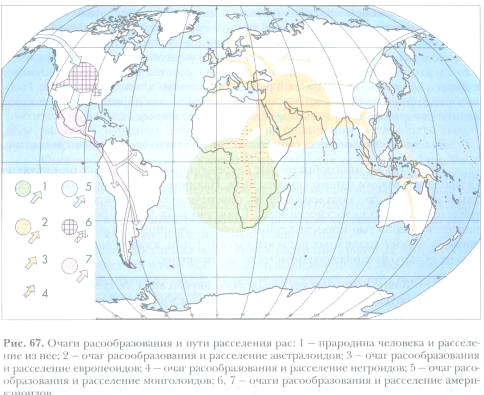
Многочисленные и очень разнообразные по составу основные расы и расовые локальные комплексы не могли возникнуть сразу на занимаемых ими обширных территориях. Не все места на планете пригодны для проживания человека, существовали также непреодолимые ранее географические препятствия для расселения в виде горных хребтов, водных пространств, лесных массивов, песчаных и ледовых пустынь. На протяжении четвертичного периода радикально изменялась природная среда. Таяние мощного ледникового панцыря после вюрмского оледенения и подъем уровня Мирового океана с образованием множества островов, разъединение континентов водными преградами, высыхание больших пространств и образование пустынь - далеко не полный сценарий возникновения изоляционных барьеров в период, когда происходила расовая дифференциация человеческого вида. Изменения морфологического облика земного шара делали невозможным одновременное и повсеместное формирование расовых комплексов в пределах занимаемого ими сейчас географического пространства. Расселение первобытного человечества было мозаичным, и, соответственно, процессы расогенеза носили локальный характер, приуроченный к определенным географическим центрам или очагам. Под географическими центрами имеются в виду области, в которых образование человеческих рас определялось в основном физическими условиями и различалось по интенсивности. Для обозначения таких областей считается более удачным слово «очаги», так как им делается акцент на динамичность расообразовательного процесса. Исторически первой и наиболее масштабной была задача определения очагов происхождения основных рас. Исходя из преобладающего расселения их локальных подразделений в настоящее время, логично предположить, что монголоидная раса формировалась в Восточной или Центральной Азии, европеоидная - в Европе, Передней Азии или сопряженных географических регионах, негроидная в - Африке, австралоидная - на архипелагах Океании. В целом этот аргумент серьезно не дискутируется в литературе еще и потому, что его невозможно абсолютно ни доказать, ни опровергнуть. Расообразование представляло собой не монотонный, плавно текущий процесс, а дискретный не только в пространстве, но и во времени. Пространственная дифференциация расовой структуры человечества, приуроченная к определенным географическим координатам, имела временную последовательность в этапах расогенеза, отраженную в их исторической древности. В.П. Алексеев выделяет четыре очага расообразования: первичный, вторичный, третичный, четвертичный, и соответствующее число этапов этого исторического процесса. Первый этап связан с возникновениям первичных очагов расогенеза, в которых сформировались три основные расы - европеоидная, негроидная, монголоидная. Он был наиболее длительным, продолжался около 200 тыс. лет, охватывая какой-то отрезок нижнего палеолита, средний палеолит и начало верхнего. Отсюда следует предположить, что разделение на основные расы началось еще до появления Homo sapiens как уже окончательно сформировавшегося вида. До сих пор сохраняются две противоположные точки зрения по вопросу о полигенетическом или моногенетическом происхождении основных рас, т.е. возникновении их от разных независимых друг от друга прародителей или из одного общего корня. Соответственно такому размежеванию расходились и представления о географических координатах расогенеза - полицентрическом или моноцентрическом. Первые представления о полигенетическом происхождении рас появились за три с половиной тысяч лет назад в трудах иранских сочинителей. Проповедники зороастризма уверяли, что человечество исходно состояло из двух основных рас северной и южной, первая имела своим горнилом мифическое «светлое» - начало, от которого произошло белое население, вторая «темное» начало, породившее негров, а также высших антропоидов шимпанзе и гориллу. В средневековой Европе адептами христианской религии в сознание людей вдалбливалась догма о божественном (по научному моногенетическом) творении человека и миф о таковом же создании человеческих рас. Еще в начале ХІХ в. для поддержания полигенизма Вирей во Франции привлекал библейскую легенду о сыновьях Ноя: родоначальником белой расы назывался Яфет, смуглой (монголоидной) Сим, чернокожей «отверженный богом» Хам. Без ссылок - на библейскую «классификацию» принцип полигенизма подкреплялся авторитетами Вольтера, Д. Юма, А. Катрфажа, Т. Гексли. Исходя из крайнего моноцентризма, согласно которому человеческий вид исторически появился на ограниченной территории, причем не существенно в Передней, Центральной или Восточной Азии, Восточной или Центральной Африке, следует признать, что и основные расы или их предковые формы возникли здесь же, в недрах ближайшего предка гомо сапиенса (пресапиенса). Новейшие данные сравнительной морфологии, палеоантропологии, археологии позволяют внести в такое утверждение некоторые существенные коррективы. Предполагается, что еще в среднем палеолите возникли два первичных географических центра, в которых независимо появились предковые формы двух расовых стволов: восточного и западного. От них произошли соответственно монголоидная раса (восточный центр), европеоидная и негроидная расы (западный центр). Мощным географическим барьером между этими центрами оказались горные массивы Центральной Азии, трудно преодолимые без специальных для этого средств и в настоящее время. Богатые коллекции ископаемых синантропов из пещеры Чжоу-коудянь по морфологическому строению, особенно широколицего черепа, указывают на генеалогическую связь с предковой формой монголоидной расы, близкой и к другим восточным археоантропам, в частности, к яванскому питекантропу. К сожалению, находки палеоантропов в Восточной Азии фрагментарны и немногочисленны, поэтому весьма затруднительно установить, какая из их групп могла быть исходным предком монголоидной расы или как-то филогенетически была связана с ней. Утверждение о первичном центре происхождения монголоидной расы в Восточной Азии подтверждается и особенностями восточной культуры палеолита. Из первичного центра родоначальники монголоидной расы широко распространились в Юго-Восточную и Северную Азию, проникли в Америку, в итоге образовались две ветви общего Амеро-Азиатского ствола: азиатская и американская с многочисленными локальными расами и группами популяций. На основании хорошо сохранившихся ископаемых останков палеоантропов в Палестине и Ираке Передняя Азия считается наиболее вероятной прародиной человека современного физического типа. Неандертальцы из пещер Схул и Кафзех имели совершенно очевидное сочетание европеоидных и негроидных признаков, без примеси явных монголоидных черт. Следовательно, с уверенностью можно заключить, что именно здесь в эпоху среднего и, возможно, уже нижнего палеолита закладывался первичный антропологический тип, из которого затем сформировалась европеоидная раса, расселившаяся по Европе, Северной Африке, Южной Азии, и негроидная раса, проникшая на территорию Центральной и Южной Африки и Океании, где она приняла дефинитивные черты современного морфотипа в географических модификациях локальных рас. Палеоантропологические материалы дополняются сравнительно-морфологическими наблюдениями над очень сходным возрастным (онтогенетическим) развитием европеоидов и негроидов. Наконец, археологические раскопки показали различие каменных технологий в восточных и западных провинциях: на востоке преобладают отщепы, пластины, односторонне обработанные, орудия (чопперы), на западе более совершенные ручные рубила. Таким образом, Передняя Азия и Восточное Средиземноморье являются наиболее вероятным первичным очагом, где сформировались общие предки европеоидной и негроидной рас. Последующее их расселение в местообитания с различными экологическими условиями и относительной изоляцией завершилось образованием множества локальных рас и групп популяций, объединяемых в три ветви общего Евро-Африканского ствола: европеоидную, африканскую, океанийскую. Если обладать способностью к восприятию научной фантастики и отрешиться на время от существующих концепций прародины человеческих рас, советуем познакомиться с оригинальной схемой рассуждений по данной теме, названной «гипотезой Гиперборей». Но прежде чем аннотировать ее, остановимся на мнении автора о полигенетическом происхождении двух главных расовых стволов, сконструированных им по принципиально иным соображениям, чем принятым в систематике. Вместо разделения всего расового состава человечества на амеро-азиатский и евро-африканский стволы, как считает этот автор, более обоснованной будет другая дивергенция и, соответственно, иное сочетание корневых слов в новых названиях - евро-американский и негроидно-монголоидный стволы. Далее им утверждается, что эти расовые стволы имели разные центры происхождения. Около 200-300 тыс. лет назад путем эволюционного процесса длительностью более 30 млн. лет в Восточной и Экваториальной Африке из Homo erectus сформировалась негроидная ветвь второго из выделенных стволов. Монголоидная ветвь этого же ствола возникла в Восточной или Центральной Азии, родоначальником которой был синантроп (Homo pekinensis), о чем свидетельствует такой, хотя и частный, но общий признак для него и монголоидов, как лопатообразные резцы. Приводятся аргументы, доказывающие, по мнению Б.О. Куценко, антропологическую близость европейской и американской рас и общность территории их происхождения, а также гораздо большее сходство их по морфотипу, чем сходство между монголоидами и американоидами. Краниологические показатели и группы крови у европейцев и коренных американцев во многом совпадают, различия же между ними легко объясняются своеобразием микроэволюции в географически отдаленных регионах. У североамериканских индейцев и у европейцев весьма близки лицевые указатели (узкое лицо, узкий удлиненный нос), доминирует первая группа крови, в то время как у монголоидов широкое лицо, относительно широкий и уплощенный нос, преобладает третья группа крови. Все эти совпадения и различия наводят на мысли не только о генеалогическом единстве, но и общем центре происхождения европейской и американской рас. В оригинальной попытке доказать историческую реальность этого центра и заключается квинтэссенция «гипотезы Гипербореи». В целом она не лишена логической стройности и аргументации, чтобы ее принимать за одну из очередных научно-фантастических выдумок. Согласно принятому на сегодня мнению, американская ветвь сформировалась от монголоидных предков, которые из Восточной или Центральной Азии мигрировали в северном направлении, через Берингов пролив проникли на территорию Нового Света и в течение тысячелетия заселили весь американский континент. В последующее время этот протоморфный монголоидный тип дивергировал на азиатскую плосколицую и узкоглазую расу и американскую разновидность с удлиненной головой и узким резко выступающим носом. Этой современной парадигме, считает Куценко, противоречат антропологические и археологические данные. О различиях между монголоидной и американоидной расами по строению черепа и группам крови выше упоминалось. По археологическим материалам (из разных публикаций) человек появился в Америке 25-40 тыс. лет назад, а находки на Аляске датируются максимум 20 тыс. лет. Кроме того, даже если допустить заселение американского континента азиатскими пришельцами, на их прародине должен был сохраниться исходный антропологический тип, формировавшийся здесь на протяжении сотен тысяч лет, а мигранты переадаптироваться к условиям этого континента. «Произошло же все с точностью до наоборот»: американские индейцы почти полностью унаследовали азиатский морфотип палеоантропов, а современное население Азии с ним рассталось. Из данного силлогизма вполне допустим вариант заселения Азии из Америки, но зададим вопрос: заселения кем? Американские мигранты в Азию не могли возникнуть в Новом Свете, так как здесь отсутствовали подходящие гоминидные предки для формирования сапиенса. Аналогичная ситуация складывалась в Европе. Наиболее древние антропологические находки Homo sapiens, отмечает автор гипотезы, обнаружены в северных ее областях (где конкретно, не указывается), а основным направлением расселения неандертальцев было движение с севера на юг. Однако северная и центральная части Европы были покрыты в то время льдом, поэтому возникают вопросы: где же находилось исходное место, из которого распространялись в южном направлении европеоидная и американоидная группы, и что вызвало такую миграцию? Ответы на данные вопросы, считает Куценко, дают палеонтологические, геологические, океанологические и климатологические исследования гипербореальной части планеты, существовавшей 250-300 тыс. лет назад. По его предположению, в это время уровень мирового океана был на 1000 метров ниже современного. Смещение магнитного и географического полюсов в верхнем плейстоцене вызвало обледенение части Северной Европы и Северной Америки. Отсутствовали Карское и Баренцево моря, на их месте и севернее до Шпицбергена простиралась равнина, на которой господствовал мягкий климат, о чем свидетельствуют остатки субтропической растительности, костехранилища мамонтов и других животных. Автор заключает: «Вот тот центр, та земля, из которой вышли европеоидная и американоидная расы. Одновременное заселение из этого региона и Азии и Америки объясняет возникшие ранее проблемы с расселением рас. Синантроп же не имеет никакого отношения к американоидной расе и является, видимо, как и erectus, переходной формой для монголоидов, имеющих, как и негроидная раса, животное происхождение». К такому выводу можно предъявить два серьезных возражения. Во-первых, постулат об эпицентре, из которого распространялись в южном направлении европеоиды и американоиды, ничем не подкреплен документально, кроме указания на существование в гипербореальной области суши с теплым климатом. Этот контраргумент можно, конечно, отвести ссылкой на то, что палеоантропологические следы погребены под толщей морской воды. Во-вторых, из рассуждений автора можно заключить, что европеоиды и американоиды исторически формировались в географически изолированных регионах, т.е. аллопатрически, правда, где первые неизвестно, вторые в Азии. В таком случае как объяснить их значительное морфологическое сходство? Расологический же по духу тезис о «животном» происхождении монголоидной и негроидной рас вообще не подлежит научному обсуждению. Слабость фактической базы в обоснование «гипотезы Гипербореи» отводит ей место более или менее вероятного предположения в объяснении проблемы стволового расогенеза, как впрочем и многих других умозрительных построениях. Итак, первый этап расообразования это возникновение первичных очагов, в которых закладывались основания для формирования расовых стволов: восточного, состоящего из азиатских монголоидов, и западного, включающего исходных предков европеоидов и монголоидов. Что касается австралоидной расы, уже давно существует точка зрения о ее гибридном происхождении от смешения монголоидной и негроидной ветвей. Принятая ранее парадигма о монголоидных предках американской расы в последнее время подвергается сомнению, но не приведены еще достаточно убедительные аргументы о других ее филогенетических корнях. Второй этап приурочен к вторичным очагам и возникновению расовых ветвей внутри основных расовых стволов. Временной отрезок его составляет порядка 15-20 тыс. лет (эпоха верхнего палеолита и начало мезолита). Формирование вторичных очагов расогенеза происходило по мере заселения новых экологических зон на территории Старого Света и, особенно, с проникновением человека в неосвоенные ранее регионы Океании, Австралии и Америки. На протяжении второго этапа создаются исходные основания для азиатской и американской ветвей восточного ствола, африканской и океанийской ветвей западного ствола. Третий этап отмечен возникновением третичных очагов расогенеза, обусловленным биологическим прогрессом человечества значительным увеличением общей численности и расширением территорни эйкумены. В абсолютных цифрах длительность его определяется в 10-12 тыс. лет, т.е. включает конец мезолита и переход к эпохе бронзы (энеолит). Для более многочисленного человеческого сообщества требовались интенсивное использование уже заселенных мест и освоение новых регионов. Оба эти процесса, особенно второй, сопровождались соответствующими морфофункциональными приспособлениями, часть которых несла на себе печать расовых различий. Уже на втором этапе расообразования зафиксированы крупные миграции населения, резко усилившиеся на третьем этапе вследствие увеличения демографического давления. Поэтому, наряду с сугубо экологическими факторами (борьба за существование в разных ее формах), нарастает роль метисации между расовыми типами, причем не только в ближнем соседстве, но и на больших расстояниях. На третьем этапе значительно обогащается расовый состав человечества в результате образования локальных рас в пределах основных расовых ветвей, заселения практически всех пригодных для проживания экологических ниш планеты. Четвертый этап характеризуется возникновением четвертичных очагов и дальнейшей расовой дифференциацией до почти полного оформления расовой структуры современного человечества. Продолжительность его - от рубежа ѴІ-Ѵ тысячелетий до начала новой эры, т.е. 3-3,5 тысячи лет, охватывая эпоху бронзы и раннего железа. На этом этапе особенно усиливается межрасовое смешение в связи с активизацией миграционных процессов, освоением новых регионов, расширением международной торговли и работорговли, масштабных военных действий. В результате возникает множество групп популяций в пределах локальных рас с устойчивым сочетанием признаков у вновь возникших расовых вариантов. К сказанному добавим, что и в последующий период истории человечества происходили заметные расогенетические события, о чем свидетельствуют данные палеоантропологии, соматологии, этнографии, общественной истории, лингвистики. Основным фактором продолжающейся расовой дифференциации в последние два тысячелетия, особенно после великих географических открытий, была интенсивная метисация между основными расами и, в особенности, между локальными их подразделениями в разных географических регионах. На островах Океании и Гавайском архипелаге образовалось население с очень сложным генетическим составом от американо-японскополинезийских браков. В Центральной Америке (Мексика, США), на Антильском архипелаге (Куба, Гаити) после заселения их испанцами, французами, англичанами, африканскими неграми сформировались локальные группы популяций, исключительно разнообразных по цвету кожи от светлого до очень темного с богатой - гаммой промежуточных вариантов. Все эти микроэволюционные последствия вызывались межрасовым смешением, полигенией, а также генетической изменчивостью, закрепляемой стабилизирующим отбором. Очень разные по окраске кожи популяции распространились по всей территории американского континента от самого юга до севера, т.е. не подчиняются экологическому правилу Глогера. По другим антропологическим особенностям (размерам тела и его частей, ширине носа) картина аналогичная, если ее рассматривать в свете других экологических правил. В Южной Америке (Бразилия, Аргентина, Чили) проживает население, говорящее на испанском и французском языках, но образовалось оно в результате метисации европейцев и негров, вместе с ассимиляцией индейских племен. Освоение Сибири представителями европеоидной расы (русские, украинцы) сопровождалось смешением их с местным населением, что отразилось на формировании расовых вариантов с включением признаков западного и восточного расовых стволов, образованием промежуточных между ними южносибирской (туранской) и уральской локальных рас. На завершающем этапе, помимо смешения, определенное значение в расовой дифференциации имели другие факторы, основательное изучение которых дело будущего. Сейчас можно сказать, что на нее влияли изменения качества питания и экологической ситуации, видимо, некоторые социальные причины. Особенности эволюции расового состава человечества от начала новой эры до настоящего момента дают основание говорить о пятом этапе расогенеза, отличающегося появлением больших контингентов смешанного населения. Таким образом, человеческая раса это не суммарное образование из однотипных индивидуумов, а группа популяций, объединяемых комплексом сходных признаков. Значительная их часть подвержена непрерывной изменчивости, которая определяет плавные переходы между расами. Другие признаки являются типичными маркерами основных рас, модифицируются в локальных расах, наследуются в комплексе или независимо. Раса представляет собой внутренне противоречивое и динамичное образование: как результат исторического развития она относительно стабильна, по многим признакам демонстрирует приспособленность к условиям своего формирования, занимает определенное географическое местоположение, а как объект эволюционного процесса неустойчива, изменчива в пространстве и во времени под действием как внешних (экологические условия, метисация), так и внутренних (дрейф генов, демографические сдвиги, изменения в динамике индивидуального развития) факторов. Уже в недрах первоначально возникающего человеческого вида (нижний и средний палеолит) закладывались элементы его расовой структуры, из которых в последующее время формировались расовые стволы, их отдельные ветви, локальные расы и группы популяций в пределах этих рас. В итоге складывалась сложная по многообразию вариантов картина расового состава народов мира, представленная в финале сейчас. Расогенез проходил в рамках микроэволюции гомо сапиенса, составляя с ней одно целое, что является ярким свидетельством реальности биологической эволюции человеческого вида в прошлом, ориентацией на его развитие в перспективе и увлекательной областью будущих исследований. Историческое формирование человеческих рас определялось тремя основными и независимыми друг от друга факторами: естественным отбором, дрейфом генов, метисацией. Два из них - отбор и дрейф генов идентичны движущим силам микроэволюции природных видов. Отбором расогенез осуществлялся на основе создания приспособлений к различным эколого-географическим условиям, дрейфом генов распространялись в микроизолятах нейтральные признаки. Метисация путем смешанных браков как фактор расогенеза не подчиняется контролю отбора, в отличие от природных видов, а полностью зависит от социальных предрасположений или запретов. Эта особенность метисации у человека обеспечивает все более возрастающую ее роль в расообразовательном процессе и вместе с тем стирает межрасовые различия. С достаточным основанием можно прогнозировать постепенное выравнивание созданного длительной эволюцией расового многообразия рода человеческого, но это весьма отдаленная перспектива, и насколько она будет для него полезной или вредной, предсказать сейчас невозможно.